Раздел 1
БИОЛОГИЧЕСКИЕ ОСНОВЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
1.3. Онтогенетический уровень
организации жизни
1.3.2. Основы генетики человека
1.3.2.12. Гены аутосом и половых хромосом
Еще в конце XIX столетия ученые
обратили внимание на различие по одной из пар гомологичных хромосом в хромосомных наборах самцов и
самок. В диплоидных соматических клетках человека хромосомы каждой из двадцати двух
пар (нестатеві хромосомы - аутосоми) сходны между собой, а хромосомы двадцать
третьей пары - разные (половые хромосомы - гетерохромосоми). Одна из
гетерохромосом - паличкоподібної формы (так называемая Х-хромосома), другая - гачкоподібна
(Y-хромосома). Итак, кариотип женщины
можно обозначить как 44А + XX, а мужчины - 44А + ХY (символом ”А ” обозначают аутосоми).
Поскольку во время мейоза гомологичные
хромосомы расходятся к разным гамет, то у одного пола возникает только один тип
гамет (гомогаметна пол), в другой - два,
которые различаются строением
половых хромосом (гетерогаметна пол).
Существуют четыре основных типа
регуляции пола половыми хромосомами (рис. 1.114):
1. ХY-тип, за которого женский пол имеет две половые XX
хромосомы и является гомозиготной (образует гаметы одного типа - X), а мужской пол
имеет мужскую половую хромосому
- Y, которая отличается от Х-хромосомы
величиной и формой. Мужской пол гетерогаметна (образует гаметы двух типов - Х и Y). Этот тип присущ млекопитающим,
насекомым и большинства двудольных покрытосеменных растений.
2. ХО-тип - женский пол имеет две
Х-хромосомы, а мужская только одну Х-хромосому. ХO-тип встречается среди насекомых
и млекопитающих.
3. ZW-rvsn. Женский пол имеет одну
женскую половую хромосому Wi вторую отличную от нее по форме и величине
половую хромосому Z. Здесь женский пол является гетерогаметною,
а мужской пол имеет две одинаковых половых хромосомы Z и является гомологичной. ZW-тип присущ некоторым рыбам, бабочкам, птицам и очень
редко встречается среди растений.
4. ZО-тип - женский пол имеет только одну Z-хромосому и
гетерогаметна, а мужская - две Z-хромосомы
и гомогаметна. Этот тип известен только у одного из видов ящерицы.
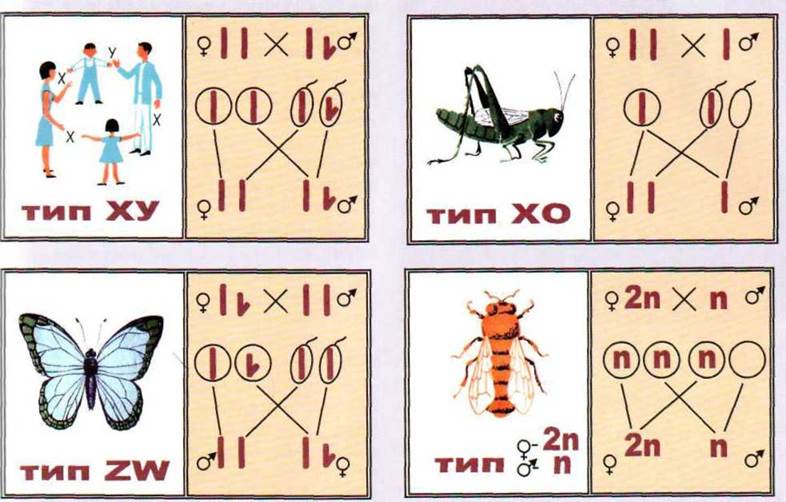
Рис. 1.114. Типы регуляции пола половыми хромосомами.
Кроме хромосомного, есть и другие
механизмы определения пола организмов. Например, у некоторых беспозвоночных животных
(коловраток, мелкого многощетинковых червей - динофілюса т.д) пол
определяется еще до момента оплодотворения. Эти животные продуцируют яйцеклетки двух
типов: большие, богатые на желток, и мелкие, с небольшим запасом питательных
веществ. Из яйцеклеток первого типа развиваются самки, а из яйцеклеток второго
- самцы.
На пол будущего организма
могут влиять факторы внешней среды. Например, у морского червя
бонелії - личинки, которые прикрепляются ко дну, развиваются в крупных (длиной
до метра) самок. Те же из них, что попадают на хоботок самки, под влиянием ее гормонов
превращаются в карликовых самцов (длиной 1-3 мм) и живут в половых
путях самки.
У общественных перепончатокрылых насекомых
(пчелы, шмели, муравьи и т.д.) самки
продуцируют два типа яиц: оплодотворенные и неоплодотворенные (партеногенетичні). Из яиц
первого типа развиваются диплоидные самки, а с другой - гаплоидные самцы.
Впоследствии в соматических клетках самцов количество хромосом удваивается, и они
становятся диплоидными.
На формирование пола некоторых рыб и
земноводных также влияют условия окружающей среды или гормоны. Во время эмбриогенеза в
них закладываются одновременно зачатки мужских и женских половых желез,
но развивается только один тип. Интересный пример такого явления можно наблюдать
у рыб-"чистильщиков", самцы которых имеют "гаремы" из нескольких
самок. После гибели самца его функции берет на себя одна из самок, у которой с
недифференцированных зачатков развиваются семенники. Изменение пола в ходе онтогенеза
наблюдают у кольчатого червя офріотроха. Молодые особи этого животного
сначала становятся самцами, а достигнув определенных размеров, превращаются в самок.
Подобные примеры известны также среди ракообразных и рыб (например, в
аквариумных рыбок-меченосцев самки с возрастом способны превращаться в самцов).
Половые хромосомы, кроме определения
пола, выполняют и другие функции, поскольку содержат гены, которые влияют на различные
системы органов. Наследование признаков и свойств, обусловленных генами,
локализованными в половых хромосомах, называется наследованием, сцепленным с
статью. Это явление было открыто Т. Морганом. Характер сцепленного с полом
наследование существенно зависит от того, в каких половых хромосомах и в которых их
участках размещены те или иные гены.
В половых хромосомах содержится одна
из крупнейших групп сцепления. В ее состав входит около 90 генов. Эти гены
делятся на три подгруппы в зависимости от того, в каких участках X - и Y-хромосом они расположены.
К первой подгруппе отнесены гены,
расположены в парном сегменте половых хромосом (в той области, которая одинакова в
X - и Y-хромосом). Такие гены частично
сцепленные с полом и остаются ограниченными Х-хромосомой только к кроссинговера
между локусом их расположение. После такого кроссинговера гены переходят из X- Y-хромосому и возникает уже наследования, характерно для генов, частичное
сцепленных с Y-хромосомой. К этой подгруппе
принадлежат много генов, сцепленных с полом. Гены, локализованные в гомологичных
участках X - и Y-хромосом. обусловливают развитие
некоторых болезней:
• пигментная ксеродерма (рис. 1.115)
- заболевания, при котором под воздействием ультрафиолетовых лучей на открытых
участках тела появляются пигментированные пятна;
• болезнь Огучи - в слое палочек и
колбочек сетчатки и пигментном эпителии возникают дегенеративные изменения;
• спастическая параплегія - слабость
в нижних конечностях в результате дегенерации нервных путей в спинном или
головном мозге;
• эпидермолиз буллезный - образование
пузырей после механической травмы кожи.
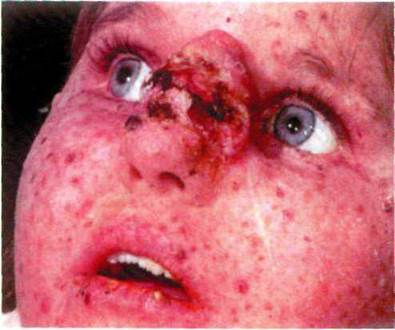
Рис. 1.115. Пигментная ксеродерма.
Вторую подгруппу составляют гены,
расположены в участке Х-хромосомы, соответствующей в которой Y-хромосомы нет. Такие гены полностью сцеплены с
статью, передаются потомкам вместе с Х-хромосомой и не могут переходить из X - в
Y-хромосому. Число генов, которые входят
в эту подгруппу, в человека достаточно велико.
Третью подгруппу составляют гены
нечетной участка Y-хромосомы. Такие гены получили название
голандричних, они ограничены только мужским полом и число их небольшое
(рыбья кожа, перепончатые пальцы, фактор развития семенников, повышенная
количество волос в наружном слуховом ходе и др.)
Признаки, сцепленные с полом, выявлено
у двудольных растений, многих животных и человека. Такие наследственные болезни, как
дальтонизм и гемофилия, вызываемые рецессивными генами, локализованными в
Х-хромосоме. Женщины, гетерозиготные по этим генами, являются носителями болезней, а у мужчин
рецессивные гены, находясь в гемізиготному состоянии, проявляют свое действие.
Есть определенные характерные особенности
наследование признаков, сцепленных с полом. Гены, локализованные в X-хромосоме,
как и при аутосомному наследовании, могут быть доминантными и рецессивными.
Отличительной чертой Х-сцепленного наследования является отсутствие передачи соответствующего
гена от отца к сыну; поскольку мужчины гемізиготні (имеют только одну
Х-хромосому), они передают свою Х-хромосому только дочерям.
Если в Х-хромосоме расположен
доминантный ген, такой тип наследования называется Х-сцепленным доминантным.
Для него характерны такие признаки:
1) если отец болен, то все его
дочери будут больными, а сыновья здоровыми;
2) больными дети будут только в
случае, когда болен один из родителей;
3) у здоровых родителей все дети
будут здоровыми;
4) заболевания регистрируется в
каждом поколении;
5) если мать больна, то вероятность
рождения зорої ребенка составляет 50 % независимо от пола;
6) болеют как мужчины, так и
женщины, но больных женщин в семье в два раза больше, чем больных мужчин.
Если в Х-хромосоме локализуется
рецессивный ген - тип наследования называется Х-сцепленным рецессивным. Для него
характерны такие признаки:
1) болеют преимущественно лица
мужского пола;
2) заболевание наблюдается у
мужчин - родственников пробанда по материнской линии;
3) сын никогда не наследует
заболевание отца;
4) если пробанд женщина, то ее отец
обязательно болен, а также больные все ее сыновья;
5) от брака больных мужчин и
здоровых женщин все дети будут здоровыми, но у дочерей могут быть больные сыновья;
6) от брака между здоровым мужчиной
и гетерозиготной женщиной вероятность рождения больного ребенка составляет 50 % для
мальчиков и 0 % для девочек;
7) если в отдельных случаях болезнь
возникает у женщины, то ее отец обязательно болен, амати является носителем.
На том основании, что отдельные признаки
встречаются чаще у представителей того или иного пола, не можно
утверждать, что это признак, сцепленная с полом. Например, склонность к
преждевременного облысения - доминантный признак, который встречается преимущественно в
мужчин; синдром Лоуренса-Мура-Бидля и одна из форм олигофрении - это
рецессивные признаки, которые чаще проявляются у мужчин, чем у женщин. Однако гены,
которые детерминируют упомянутые признаки, локализованы не в половых хромосомах, а в
аутосомах. Неодинаковое распределение признаков между представителями разных полов
обусловлен тем, что женский генотип создает среду, которая тормозит или ослабляет
проявление этих генов. Обычно говорят, что наследование таких признаков ограничивается или
контролируется статью.
Особь женского пола может быть как
гомо-, так и гетерозиготной по генам, которые локализованы в Х-хромосоме, а рецессивные аллели генов в нее
проявляются только в гомозиготном состоянии. Поскольку у лиц мужского пола
только одна Х-хромосома, все локализованные в ней гены, даже рецессивные,
проявляются в фенотипе. Такой организм называют гемізиготним.