Раздел 1
БИОЛОГИЧЕСКИЕ ОСНОВЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
1.3. Онтогенетический уровень
организации жизни
1.3.2. Основы генетики человека
1.3.2.9. Сцепленное наследование генов
Доказано, что количество наследственных
признаков организма значительно превышает число хромосом гаплоидного набора. Так, в
гаплоидном наборе классического объекта генетических исследований - мухи-дрозофилы -
есть только четыре хромосомы, но число наследственных признаков и, соответственно, генов, которые их
определяют, несомненно, значительно больше. Это означает, что в каждой хромосоме
находится много генов. Поэтому вместе с признаками, которые наследуются независимо,
должны существовать и такие, которые наследуются сцеплено друг с другом, ибо они определяются генами, расположенными в одной
хромосоме. Такие гены образуют группу сцепления. Количество групп сцепления в
организмов определенного вида равно числу хромосом в гаплоидном наборе
(например, у дрозофилы 1п=4, у человека 1п=23).
Экспериментальные исследования явления
сцепленного наследования провел выдающийся американский генетик Т. X. Морган со
своими сотрудниками. Их результаты обосновали предложенную ими
хромосомную теорию наследственности.
Следует отметить, что Т. X. Морган, как
в свое время Г. Мендель, удачно выбрал для исследований муху-дрозофилу, которая впоследствии
стала классическим объектом для генетических экспериментов. Дрозофил легко
содержать в лабораториях, они имеют значительную плодовитость, быструю смену поколений
(при оптимальных условиях содержания новое поколение возникает каждые полторы-две недели),
небольшое число хромосом, что упрощает наблюдения.
Явление сцепленного наследования Т. X.
Морган установил в таком опыте. Самцов дрозофилы, гомозиготных по
доминантными аллелями окраски тела (серая) и формой крыльев (нормальные), зхрестили
с самками, гомозиготными по соответствующим
рецессивными аллелями (черное тело - недоразвитые крылья). Генотипы этих особей
обозначили соответственно EEVV и eevv. Все гибриды первого поколения имели
серое тело и нормальные крылья, то есть были гетерозиготными по обоим парами апелів
(генотип - EeVv) (рис. 1.109).
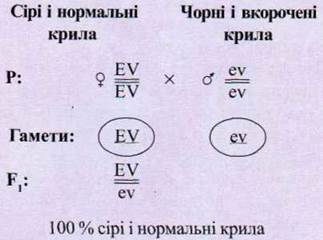
Рис. 1.109. Схема скрещивания при
полном сцеплении.
Затем гибридов зхрестили с
особями, гомозиготными по соответствующим рецессивными аллелями (анализирующее
скрещивание).
Теоретически можно было ожидать два
варианты расщепления. Если бы гены, которые обусловливают окраску тела и форму крыльев,
содержались в негомологичних хромосомах, то есть наследовались независимо, расщепление должно быть таким: 25 %
особей с серым телом и нормальными крыльями, 25 % - с серым телом и
недоразвитыми крыльями, еще 25 % - с черным телом и нормальными крыльями и 25 %
- с черным телом и недоразвитыми крыльями (то есть в соотношении- 1:1:1:1).
Если бы эти гены размещались в одной хромосоме и наследовались сцеплено, то
было бы получено 50 % особей с серым телом и нормальными крыльями и 50 % - с
черным телом и недоразвитыми крыльями (то есть 1:1).
На самом деле 41,5 % особей имели серое
тело и нормальные крылья, 41,5 % - черное тело и недоразвитые крылья, 8,5 % - серое
тело и недоразвитые крылья и 8,5 % - черное тело и нормальные крылья, т.е.
расщепление приближался к соотношение фенотипов 1:1 (как в случае сцепленного
наследования), но вместе с тем проявились все четыре варианта фенотипа (как в
случае независимого наследования) (рис. 1.110).

Рис. 1.110. Схема скрещивания при
неполном сцеплении.
На основании этих данных Т. X. Морган
предположил, что гены, определяющие окраску тела и форму крыльев, расположенные в
одной хромосоме, но в процессе мейоза при образовании гамет гомологичные
хромосомы могут обмениваться участками, то есть имеет место явление, получившее
название перекрест хромосом, или кроссинговер.
Кроссинговер (от англ. crossing-over - перекрест) - обмен участками гомологичных хромосом в
процессе клеточного деления, преимущественно в профазе первого мейотического деления, иногда
в мітозі.
Опытами Т. Моргана, К. Бриджеса и
А. Стертеванта было показано, что нет абсолютно полного сцепления генов, при
которого гены передавались бы всегда вместе. Вероятность того, что два гена,
локализованные в одной хромосоме, не разойдутся в процессе мейоза, колеблется в
пределах 1-0,5. В природе преобладает неполное сцепление, обусловленное перекрестком
гомологичных хромосом и рекомбинацией генов (рис. 1.111).
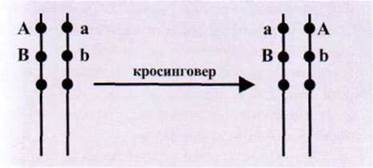
Рис. 1.111. Пример рекомбинации
генов в хромосомах вследствие кроссинговера.
Цитологическая картина кроссинговера
была впервые описана датским ученым Ф. Янсенсом.
Кроссинговер проявляется только
тогда, когда гены находятся в гетерозиготном состоянии (AB/ab). Если гены находятся в гомозиготном состоянии (AB/AB или ab/ab), обмен идентичными участками не дает
новых комбинаций генов в гаметах и в поколении.
Частота (процент) перекрестка между
генами зависит от расстояния между ними: чем дальше они расположены друг от
друга, тем чаще происходит кроссинговер. Т. Морган предложил расстояние
между генами измерять кроссинговером в процентах, по формуле:
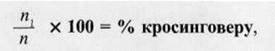
где n - общее число особей в F;
n1 - суммарное число кроссоверных особей.
Отрезок хромосомы, на котором
осуществляется 1 % кроссинговера, равна одной морганиде (условная мера расстояния
между генами). Частоту кроссинговера используют для того, чтобы определить
взаимное расположение генов и расстояние между ними. Для построения генетической карты
человека пользуются новыми технологиями. Построены цитогенетические карты
хромосом.
Различают несколько типов
кроссинговера: двойной, множественный (сложный), неправильный, неровный.
Кроссинговер приводит к новому
сочетание генов, что вызывает изменение фенотипа. Кроме того, он наряду с мутациями есть
важным фактором эволюции организмов.
Указанные доказательства были положены в
основу хромосомной теории наследственности:
1. Гены размещаются в хромосомах по
длине в линейном порядке; различные хромосомы содержат неодинаковое число генов;
набор генов каждой из негомологичних хромосом уникален.
2. Аллельные гены занимают определенные и
идентичные локусы (места) гомологичных хромосом.
3. Гены, расположенные в одной
хромосоме, образуют группу сцепления, благодаря чему имеет место сцепление некоторых
признаков, которые вместе (сцеплено) передаются потомкам. Количество групп сцепления
равно гаплоидному набору хромосом. Сцепление не абсолютное.
4. Во время мейоза, который происходит
только при образовании гамет, диплоидные число хромосом уменьшается вдвое
(гаплоидные число). Это соответствует закону расщепления, согласно которому генетический
материал обоих родителей по-разному комбинируется в гаметах.
5. Согласно закону независимого
распределения отцовские и материнские наборы не сцепленных генов расщепляются
независимо друг от друга. Если не - сцепленные гены расположены в разных
хромосомах, то во время мейоза материнские и отцовские хромосомы должны
распределиться между гаметы случайно.
6. Между генами гомологичных
отцовских и материнских групп сцепления могут происходить, благодаря
кроссинговера, реципрокные рекомбинации. Этому соответствует образование хиазм под
время конъюгации гомологичных хромосом в мейозе (генетический кроссинговер).
7. Сила сцепления между генами
обратно пропорциональна расстоянию между ними. Чем ближе расположены гены в одной
хромосоме, тем сильнее их сцепление, тем меньше будет возникать рекомбинаций между
ними, и наоборот. Расстояние между генами измеряется в процентах кроссинговера. Один
процент кроссинговера соответствует одной морганиди.
8. Каждый биологический вид
характеризуется специфическим набором хромосом - кариотипом.