Раздел 1
БИОЛОГИЧЕСКИЕ ОСНОВЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
1.3. Онтогенетический уровень
организации жизни
1.3.1.7. Оплодотворение
Половое размножение животных и растений
сопровождается оплодотворением - слиянием двух гамет: мужской и женской. В
результате образуется оплодотворенная яйцеклетка - зигота, которая дает начало
развития нового поколения организмов. Только в 1875 году было доказано, что в
основе процесса оплодотворения лежит слияние ядер одной женской и одной мужской
половых клеток, и тем самым объединения их хромосом.
В результате объединения гаплоїдних наборов хромосом восстанавливается диплоидные их число.
Существуют два способа оплодотворения:
внешнее и внутреннее. Почти все водные позвоночные животные (рыбы, земноводные и
др.) откладывают икру (яйцеклетки) и сперму в воду, где и происходит
оплодотворения. Исключение составляют водные млекопитающие (ластоногие, китообразные), живородящие
рыбы и некоторые земноводные, которым свойственно внутреннее оплодотворение.
У наземных животных оплодотворение
происходит в половой системе самки, и зародыш развивается или в середине ее
тела (внутриутробное развитие у млекопитающих), или в яйцах, покрытых скорлупой
(насекомые, пресмыкающиеся, птицы, яйцекладні млекопитающие). Яйца согреваются теплом матери,
солнечными лучами или теплом, которое выделяется при гниении органических остатков.
Зародыш развивается вне тела самки, яйцеклетка содержит большие запасы
питательных веществ, которые обеспечивают развитие зародыша.
При внутриутробном развитии
плода яйцеклетка содержит минимальное количество питательного материала.
Процесс оплодотворения у человека.
Оплодотворение может наступить сразу после завершения яйцеклеткой стадии созревания.
В этот период она покрыта слоем фолликулярных клеток, содержит гаплоидный набор -
23 хромосомы.
Во время оплодотворения осуществляются два
важные процессы: активация яйца, то есть возбуждение к развитию, и синкаріогамія,
т.е. образование диплоидного ядра зиготы в результате слияния ядер гаплоїдних
половых клеток, которые несут генетическую информацию двух родительских организмов.
Встрече гамет способствует то, что
яйцеклетки выделяют в окружающую среду химические вещества, которые активизируют
сперматозоони. Возможно, что такие вещества выделяются и клетками женских
половых путей.
Проникновение сперматозоона
происходит благодаря акросомній реакции, которая выявлена с помощью электронной
микроскопии. Расположена на переднем конце сперматозоона акросомна участок,
окруженная мембраной. При контакте с яйцом оболочка акросомы разрушается, из нее
выбрасывается акросомна нитка. выделяются ферменты: один растворяет обо лонку
яйцеклетки, и второй фермент-гиалуронидаза, который растворяет фолликулярные клетки,
окружающие яйцеклетку (рис. 1.92). Акросомна нить проникает сквозь растворимый
зону яйцевых оболочек и сливается с мембраной яйцеклетки. В этом месте с
цитоплазмы яйцеклетки образуется воспринимающий бугорок. Он захватывает ядро,
центріолі и митохондрии сперматозоона и втягивает их внутрь яйца. Плазматическая
мембрана сперматозоона монтируется в поверхностную мембрану яйца, образует
мозаичную внешнюю мембрану зиготы.
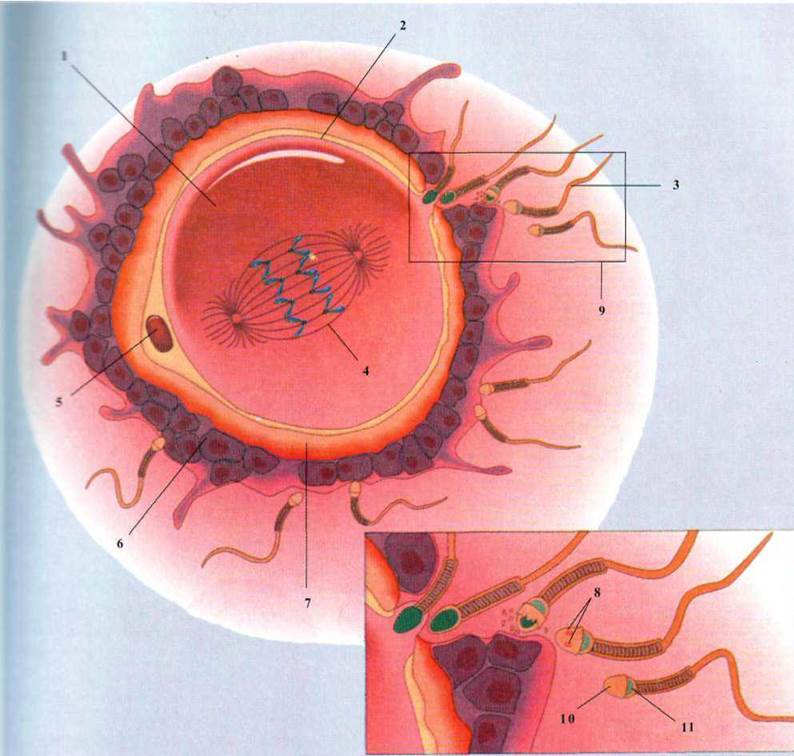
Рис. 1.92. Схема оплодотворения:
1 - цитоплазма; 2 - клеточная
мембрана; 3 - сперматозоони; 4 - мейоз II; 5 - первичный полоцит; 6 -
лучистая корона; 7 - блестящая зона; 8 - перфорации в стенке акросомы;
9 - акросомальная реакция; 10 -
акросома; 11 - ядро.
Проникновение в сперматозоона
яйцеклетку, изменяет ее обмен веществ, показателем чего является ряд морфологических и
физиологических преобразований. Повышается проницаемость клеточной мембраны,
усиливается поглощение из окружающей среды фосфора и калия, выделяется
кальций, увеличивается обмен углеводов, активируется синтез белка. Меняются
коллоидные свойства цитоплазмы. Вязкость увеличивается в 6-8 раз. В
внешнем слое яйца изменяется эластичность и оптические свойства. На
поверхности отслаивается оболочка оплодотворения; между ней и поверхностью яйца
образуется заполненный жидкостью пространство. Под ним формируется оболочка, которая
обеспечивает скрепление клеток, которые возникают вследствие дробление яйца. После образования
оболочки оплодотворения, другие сперматозоони уже не могут проникнуть в
яйцеклетки.
Показателем изменения обмена веществ и
то, что у некоторых видов животных созревания яйца заканчивается только после
проникновения в него сперматозоона. Сперматозоони проникают в яйцеклетки, когда
находятся еще на стадии созревания. Первичный полоцит выделяется по 10 часов,
вторичный - только за сутки после проникновения сиерматозоона.
Кульминационным моментом в процессе
оплодотворение-слияние ядер (рис. 1.93). Ядро сперматозоона (мужской пронуклеус)
в цитоплазме яйца отекает и достигает размеров ядра яйцеклетки (женского пронуклеуса).
Одновременно мужской пронуклеус поворачивается на 180° и центросомою вперед
движется в сторону женского пронуклеуса; последний также смещается ему навстречу.
После встречи ядра сливаются. Вследствие синкаріогамії восстанавливается диплоидный
набор хромосом. После образования синсаріона яйцо начинает дробиться.
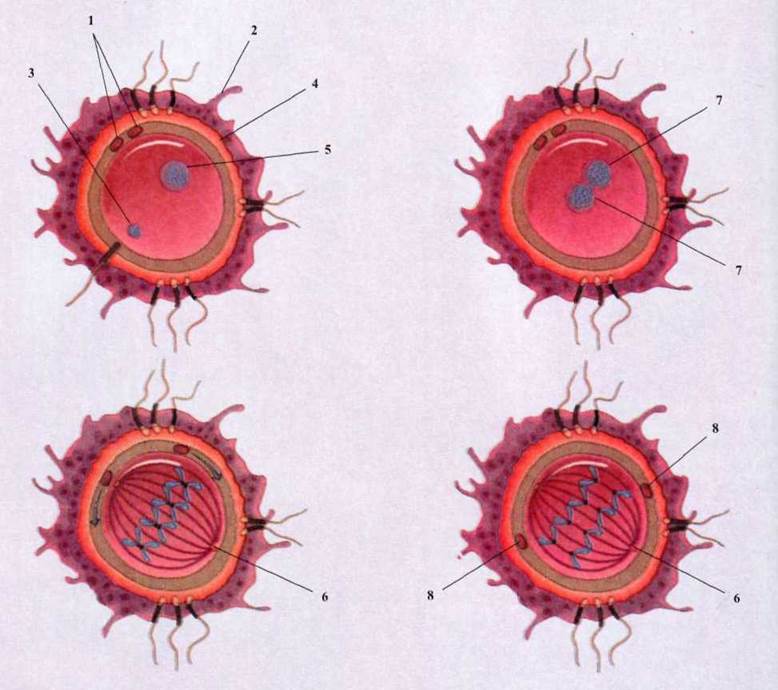
Рис. 1.93. Схема оплодотворения:
1 - первичный и вторичный полоцити;
2 - лучистая корона; 3 - ядро сперматозоона; 4 - блестящая зона; 5 - ядро
яйцеклетки; 6 - центріоля; 7 - ядра сперматозоона и яйцеклетки реплицируют
собственные ДНК; 8 - полоцити.
Изучение физиологии оплодотворения
позволяет понять роль большого количества сперматозоонів, которые принимают участие в
оплодотворении. Установлено, что когда при искусственном осеменении кроликов в семенной
жидкости содержится менее 1000 сперматозоонів, оплодотворения не происходит.
Не происходит оплодотворения и при
введении очень большого количества сперматозоонів (более 100 млн.). Это объясняется
в первом случае недостаточным количеством, а во втором - избытком ферментов,
которые необходимы для проникновения сперматозоонів в яйцеклетку.
Разработаны методики искусственного
осеменение яйцеклетки человека вне организма, и в основном это осуществляется за
медицинскими показаниями. Накануне овуляции яйцеклетку хирургическим путем вынимают
из яичника. ЕЕ помещают в специально разработанное химическое среду со
сперматозоонами, где и происходит слияние половых клеток. Зародыш на стадии
8-16 бластомеров имплантируют в матку женщины, где он и развивается.
В яйцеклетку проникает обычно один
сперматозоон (моноспермія). У насекомых, рыб, птиц и некоторых других животных в
цитоплазму яйцеклетки их может попасть несколько (поліспермія). Роль
поліспермії не совсем понятна, однако установлено, что ядро только одного из
сперматозоонів (мужской проинуклеус) сливается с женским пронуклеусом. Следовательно,
в передаче наследственной информации участвует только этот сперматозоон. Ядра
других разрушаются.
Таким образом, биологическая суть
оплодотворение полягає в следующем:
1) в результате объединения гаплоїдних
наборов хромосом восстанавливается диплоидные число
хромосом;
2) оплодотворение обеспечивает
непрерывность материального связи между поколениями организмов;
3) в результате сочетания наследственных
особенностей двух организмов у потомков образуются новые признаки, - появляется
материал для отбора, повышается изменчивость потомства, растет комбинативная
изменчивость;
4) избирательность оплодотворения
(оплодотворение только в пределах вида) обеспечивает сохранение вида как целого.
Нарушения оплодотворения, его
последствия.
Оплодотворение - одна из звеньев
биологического существования вида. Этому предшествует длительная и сложная подготовка двух
особей, во время которой они подвергаются разнообразной действия окружающей среды,
что может негативно повлиять на процесс оплодотворения.
Яйцеклетка и сперматозоид имеют
ограниченную продолжительность жизни и еще меньшую продолжительность способности к оплодотворению. Так,
у млекопитающих, и у человека в частности, освобождена из яичника яйцеклетка сохраняет
способность к оплодотворению в течение 24 ч. Нарушение этого срока временного
неизбежно приведет к потере способности к оплодотворению.
Сперматозоиды мужчины в половых
путях женщины остаются подвижными более 4 суток, но запліднювальну способность
они теряют уже через 1-2 суток. С увеличением продолжительности во времени незащищенные
клетки испытывают негативного влияния различных факторов. Последние могут вызвать
нарушения восходящего состояния генофонда гамет, что неизбежно приведет к
незапрограмованих отклонений развития зиготы с соответствующими последствиями для вида
в целом.
Скорость движения сперматозоидов, за
обычных условиях, составляет 1,5-3 мм/мин. Разное отклонение от такого поступательного
перемещение вызывает потерю способности к оплодотворению. К этому приводит также
изменение рН среды влагалища, воспалительные явления и др.
В эякуляте мужчины в среднем
содержится 350 млн. сперматозоидов, способных к оплодотворению. Если количество
сперматозоидов меньше, чем 150 млн. (или меньше 60 млн. в 1 мл), то вероятность
оплодотворения резко снижается. Итак, чрезмерная концентрация сперматозоидов в
эякуляте имеет исключительное значение в механизме оплодотворения.
Нарушения оплодотворения возникает при
патологических изменениях морфологии сперматозоидов (рис. 1.94).
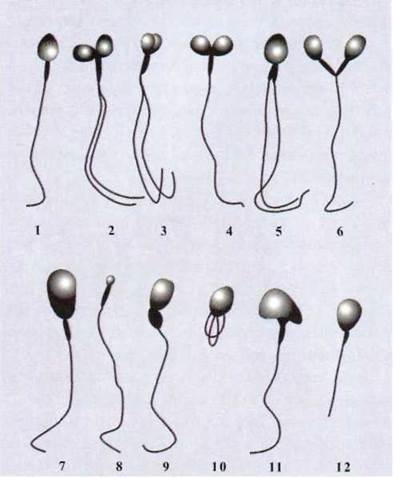
Рис. 1.94. Различные формы нарушения строения
сперматозоидов человека (2-12). Для сравнения показано сперматозоид в норме (1):
1,9 - за Максимовым; 2-5,7,11 - за Борманом; 6,8,10,12 - за Менхом и Холтом (по Пелен, 1959).
На биологическую полноценность гамет
существенно влияет срок пребывания их в половых путях женщины. Так,
перезревание сперматозоидов и яйцеклеток в женском половом тракте при различных
причин обусловливает рост частоты хромосомных аберраций в абортированных плодах (Thibault, 1972).