Раздел 1
БИОЛОГИЧЕСКИЕ ОСНОВЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
1.3. Онтогенетический уровень
организации жизни
1.3.1.5. Мейоз
Организмы, которые размножаются
половым путем, образуют половые клетки, или
гаметы. Этому предшествует особый способ деления клеточного ядра их предшественников -
мейоз (от греч. μείωσις - уменьшение). С помощью мейоза
образуются и созревают половые клетки (сперматозоиды и яйцеклетки).
Мейотичний разделение впервые описан в 1888 г. Он лежигь в основе редукции числа
хромосом (уменьшены вдвое): 2п → п. Из диплоидных клеток образуются
гаплоидные.
Если бы половые клетки содержали
диплоидный набор хромосом, то их число удваивалось бы в каждом поколении.
Поскольку каждый вид из поколения в поколение сохраняет постоянное количество хромосом,
то очевидна необходимость существования определенных механизмов, в результате которых число
хромосом уменьшалось вдвое. Это и обеспечивается редукционным делением, или
мейозом. Поскольку при оплодотворении объединяются материнский и отцовский
наборы хромосом, уменьшение их числа вдвое при образовании гамет - биологически
необходимый процесс. У животных мейоз проходит при образовании гамет, а у цветковых растений - раньше: при образовании пыльцевых зерен и
зародышевых мешков. В мейоз вступают незрелые половые клетки, которые достигают определенного дифференцировки.
Мейоз состоит из двух быстрых в
времени последовательных делений клеток: первого и второго, причем удвоение ДНК
происходит только перед первым делением. Один из них называется редукционным,
или первым мейотичним разделением, при котором число хромосом уменьшается в два раза;
другой - екваційний (ровный), или второй редукционное деление, которое напоминает
митотическое деление.
В мейоз, как и в митоз вступают
клетки с хромосомами, которые состоят из двух сестринских
хроматид. После первого деления быстро наступает второй раздел, без подготовки и без
синтеза ДНК. Второй мейотичний разделение происходит по типу митоза, только с
тем отличием, что на всех фазах будет вдвое меньшее число хромосом.
В каждом делении мейоза различают
профазу, метафазу, анафазу и телофазу. Фазы первого деления обозначают римской
цифрой И (профаза
И, метафаза И и т.д.), а фазы второго раздела цифрой II
(профаза II, метафаза II и др.) (рис. 1.89).
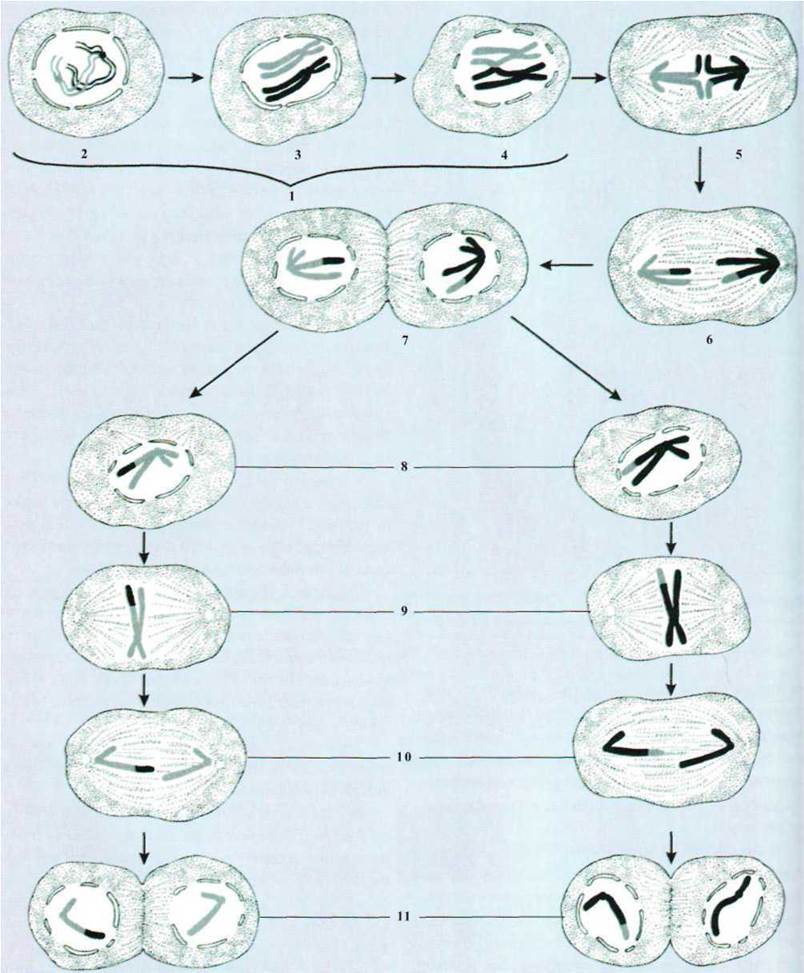
Рис. 1.89. Схема мейоза:
1 - профаза; 2 - лептонема; 3 -
зигонема; 4 - диплонема; 5 - метафаза; 6 - анафаза; 7 - телофаза I; 8 -
профаза II; 9 - метафаза II; 10 - анафаза II; 11 - телофаза II.
Профаза И. В отличие от митоза, где
каждая отдельная хромосома ведет себя независимо от других и не влияет на их
поведение, в профазе И мейоза гомологичные хромосомы объединяются, формируют
парные образования. Это длительная и сложная фаза, она характеризуется определенными
последовательными стадиями в зависимости от состояния хромосом.
Лептонема, или стадия тонких нитей.
Хромосомы становятся заметными в виде тонких нитей, количество их диплоїдна.
Зигонема - гомологичные хромосомы
сближаются попарно, образуют біваленти. Число их вдвое меньше, чем исходная
количество хромосом. Взаимное притяжение хромосом получило название конъюгация или
синапсис. Конъюгация происходит очень точно, хромосомы соединяются концами или
по всей длине. Причем сближаются каждый хромомер и каждый участок одной
гомологической нити с соответствующим хромомером и участком другой гомологичной
нити.
Пахінема, или стадия толстых нитей.
Процесс конъюгации гомологичных хромосом полностью завершается. Они настолько
сближены, что их легко можно принять за одну. Каждая хромосома в біваленті
удвоена и состоит из двух сестринских хроматид. Біваленти иногда называют тетрадами.
На стадии пахінеми происходит кроссинговер - обмен идентичными участками между
гомологичными хромосомами.
Диплонема, или стадия двойных
нитей. Хромосомы, которые образовали біваленти, начинают постепенно
отталкиваться друг от друга, оставаясь соединенными между собой в отдельных
участках (хіазмах). Каждая хромосома состоит из двух хроматид, а каждый
бівалент образует тетраду.
Переплетены одна
вокруг одной хромосомы (біваленти) постепенно раскручиваются и уменьшается число
хиазм.
Диакинез - заключительная стадия профази
И. В діакінезі біваленти резко укорочены, утолщены дочерние хроматиды каждой
хромосомы мало заметны. Хиазмы постепенно смещаются на конце хромосом.
Завершается профаза И разрушением ядерной оболочки и формированием
ахроматинового веретена.
Метафаза И. Число бивалентов вдвое
меньше диплоидного набора хромосом. Біваленти значительно короче, чем хромосомы
в метафазе соматического митоза, и размещаются в экваториальной плоскости.
Центромеры хромосом соединяются с нитями фигуры веретена. В эту фазу мейоза
можно подсчитать количество хромосом.
Анафаза И. К противоположным полюсам
веретена расходятся гомологичные хромосомы. Каждая из них состоит из двух
дочерних хроматид, соединенных своими центромерами. В этом заключается существенная
віддмінність от анафазы митоза.
Телофаза И. Начинается, когда
анафазні хромосомы достигли полюсов клетки, на каждом из них находится
гаплоидные число хромосом. Характеризуется появлением ядерной мембраны и
восстановлением структур ядра. Образуются две дочерние клетки.
Интерфаза между i И II делением мейоза
бывает очень короткой. В отличие от обычной интерфазы здесь зідсутня
репродукция хромосом. Мейоз II происходит по типу обычного митоза.
Профаза II. Эта стадия непродолжительна,
хромосомы хорошо заметны.
Метафаза II. Четко определена
двойная структура хромосом и значительная степень их
спіралізації.
Анафаза II. Происходит расхождение
удвоенных центромер, в результате чего дочерние хроматиды движутся в разных
полюсов.
Телофаза II. Завершается образованием
четырех клеток с гаплоїдним набором хромосом.
Для облегчения восприятия поведения
хромосом при мейозе и мітозі приведена сравнительная характеристика их фаз (табл.
1.11).
Таблица 1.11.
Сравнительная характеристика фаз мейоза (И) И мігозу
|
Фазы
|
Мейоз (!)
|
Митоз
|
|
Профаза
|
Более
длительная и более сложная. Происходит сближение гомологичных хромосом, образование
пар (конъюгация) и кроссинговер. Это приводит к обмену гомологичными
локусами.
|
Гомологичные
хромосомы не сближаются, пар не образуют. В конце профази они
спіралізовані и заметны, напоминают рыхлый клубок (у человека 46 хромосом).
|
|
Метафаза
|
В плоскости
экватора расположены пары гомологичных хромосом. Материнские и отцовские
хромосомы попарно расположены произвольно и со стороны одного пслюса может быть
больше родительских, а с другой - больше материнских хромосом, т.е. пары
гомологичных хромосом комбинируются случайно, а вместе с ними комбинируются и
гены, локализованные в хромосомах. Количество вариантов за счет свободного
комбинирование хромосом у человека свыше 8 млн. (223=8388608).
|
В плоскости
экватора хромосомы расположены свободно, не парами.
|
|
Анафаза
|
К полюсам
отходят целые гомологичные хромосомы, состоящие из двух хроматид, и на
полюсах размещаются по 23 хромосомы.
|
К
полюсам хроматиды отходят (у человека 46 хроматид на каждом из полюсов).
|
|
Телофаза
|
Хромосомы
с различимых в световой микроскоп (вследствие деспіралізації) могут стать
невидимыми, формируются дочерние ядра,
цитоплазма
разделяется и образуются две дочерние клетки с гаплоїдним набором хромосом,
но они состоят еще из двух хроматид.
|
С
коротких и толстых хромосомы в результате деспіралізації становятся тонкими,
длинными, невидимыми. Формируются дочерние ядра, происходит разделение цитоплазмы и
образуются две дочерние клетки с точно таким
же набором хромосом, какой был в материнской
клетке.
|
Механизмы, которые приводят к
генетического разнообразия гамет.
В процессе мейоза создаются
возможности возникновения в гаметах новых генных комбинаций.
Механизмы, которые обеспечивают
генетическую изменчивость:
1. Уменьшение числа хромосом от
диплоидного до гаплоидного сопровождается расхождением аллелей так, что каждая
гамета содержит только один аллель в локусе. Реципрокный обмен генами между хроматидами
гомологичных хромосом может происходить в профазе И мейоза. Таким образом
образуются новые группы сцепления, и соответственно генетические рекомбинации аллелей.
2. Біваленти в экваториальной плоскости
веретена деления в метафазе i И хромосом в метафазе II располагаются произвольно
и случайно. Следующее их разделение (сегрегация) в анафазах i И II соответственно
создает новые комбинации аллелей в гаметах. Такой независимый распределение приводит
к большому количеству различных хромосомных комбинаций, до случайного распределения
материнских и отцовских хромосом между дочерними ядрами.
3. Образование хиазм между
гомологичными хромосомами в профазе i И следующий кроссинговер
предопределяют новые комбинации аллелей в хромосомах половых клеток.
Генетическое значение мейотического
разделения заключается в следующем:
1. В результате мейоза каждая
материнская клетка дает начало четырем клеткам с "редукционным",
то есть уменьшенным вдвое числом хромосом.
2. Мейоз является механизмом, который
поддерживает видовое постоянство числа хромосом и обусловливает постоянство видов на
Земли. Если бы число хромосом не уменьшалось, то в каждом следующем поколении
происходило бы рост их вдвое (у родителей - 46, у детей - 92, у внуков -184,
у правнуков - 368 и т.д.)
3. Мейоз обеспечивает благодаря
случайной комбинации материнских и отцовских хромосом генетическую
разнородность гамет. То есть мейоз способствует комбінативній изменчивости (гены родителей
комбинируются, в результате чего у детей могут появляться признаки, которых не было в
родителей). Комбинативная изменчивость обеспечивает большое разнообразие человечества и
дает возможность приспособиться к изменению условий среды, способствует выживанию вида.
4. Мейоз обеспечивает разнородность
гамет по генетическим составом, способствует вследствие рекомбинации участками гомологичных
(парных) родительских хромосом образованию хромосом нового генетического состава. В
профази этому способствовал кроссинговер, в метафазе - свободное перекомбинування хромосом. То есть возникает рекомбинация
родительских наборов хромосом.