Обмен веществ в клетке
Обмен веществ (метаболизм) - совокупность химических превращений, происходящих в клетке и обеспечивающих ее рост, жизнедеятельность и воспроизведение. Обмен веществ живой клетки состоит из двух противоположно направленных видов реакций - катаболических и анаболических. Совокупность реакций распада органических соединений называется катаболизмом, или энергетическим обменом. Совокупность реакций синтеза органических соединений называется анаболізмом, или пластическим обменом. Во время расщепления (катаболизма) органических соединений (белков, жиров, углеводов) выделяется энергия, которая аккумулируется в химических связях молекул АТФ. Эта энергия используется клеткой в анаболических процессах - синтезе собственных, необходимых на данный момент времени белков, жиров и углеводов. Таким образом, энергетический и пластический обмен тесно связаны между собой потоками вещества и энергии.
По способу добывания энергии живые организмы делятся на автотрофів и гетеротрофів. Клетки гетеротрофів (животные, грибы, большинство бактерий, паразитические растения) для построения собственных биополимеров используют мономеры поглощенных и расщепленных ими органических соединений и энергию, что выделяется при этом. Автотрофи способны синтезировать собственные биополимеры из воды, углекислого газа и минеральных солей. В зависимости от источника используемой энергии автотрофів разделяют на фототрофів (аккумулируют солнечную энергию - зеленые растения, некоторые бактерии) и хемотрофів (аккумулируют энергию, выделяющуюся в результате окислительно-восстановительных процессов - сіркобактерії, нітрифікуючі бактерии).
За чувствительностью к концентрации кислорода в окружающей среде все организмы делятся на аэробов и анаэробов. Аэробы могут существовать только при достаточном содержании кислорода. Анаэробы могут быть облигатными (могут существовать только в бескислородных условиях) или факультативными (способны к жизни в широком диапазоне концентраций кислорода).
Обмен углеводов
В живых клетках глюкоза или полимеризуется в гликоген (крахмал), который служит запасным питательным материалом, или, расщепляется с выделением энергии.
Энергетический обмен углеводов
В организме гетеротрофів подвергаются ферментативному расщеплению углеводы, поглощенные из окружающей среды. В организме автотрофів расщеплению подвергаются углеводы (крахмал), синтезированы из неорганических веществ.
Дыхательную цепь митохондрий состоит из нескольких белковых комплексов, расположенных во внутренней мембране митохондрий и способных улавливать и транспортировать электроны. Согласно хеміосмотичною гипотезе Митчелла транспорт электронов по дыхательной цепи сопряжен с переносом протонов (Н
+) из матрикса митохондрий в міжмембранний пространство. В результате этого с обеих сторон внутренней мембраны возникает разность концентраций Н
+, так что протоны пытаются вернуться в матрикс. Этот процесс осуществляется с помощью специфических белков, которые пронизывают внутреннюю мембрану и формируют канал. Транспорт протонов через этот канал связан с синтезом АТФ.
Белки дыхательной цепи обнаружены во всех клетках еукаріотичних организмов (животных, растений, грибов) и в клетках аэробных прокаріот.
Благодаря процессу окислительного фосфорилирования аэробный катаболизм углеводов, оказывается гораздо эффективнее анаэробный, позволяя усвоить до 40 % энергии сгорания глюкозы. Часть энергии, которая запасается в форме АТФ, освобождается в виде теплоты и используется в теплокровных животных для поддержания постоянной температуры тела.
Пластический обмен углеводов в гетеротрофных организмов
Моносахариды, поступившие в цитоплазму, могут не только подвергаться расщеплению с выделением энергии, но и служат материалом для синтеза собственных биополимеров клетки. Глюкоза с помощью специфических ферментов полимеризуется с образованием гликогена (этот процесс называется гликогеноз). При этом расходуется энергия АТФ. Синтезированный гликоген накапливается в цитозоле в виде гранул и есть запасной питательным веществом. При необходимости он окисляется до глюкозы, которая включается в гликолиз. Глюкоза, образовавшаяся в результате распада гликогена в клетках печени млекопитающих выходит в кровь и является источником энергии для нейронов и мышц. Поскольку большинство реакций гликолиза являются обратимыми, клетка способна синтезировать глюкозу из других соединений - например, ацетил-КоА (этот процесс называется глюконеогенезом).
Пластический обмен углеводов в фототрофных организмов - фотосинтез
Фотосинтез - это процесс преобразования энергии солнечного света в энергию химических связей и синтеза органических соединений (углеводов) из неорганических (вода и углекислый газ).
Основным фотосинтетичним пигментом высших растений является хлорофилл. Процесс фотосинтеза состоит из двух взаимосвязанных этапов - световой и темновой фаз. Световая фаза проходит только при наличии света, с помощью фотосинтетических пигментов в тилакоїдах хлоропластов. Реакции темновой фазы не требуют для своего осуществления света и происходят в строме хлоропластов. Суммарные уравнения световой и темновой фаз:
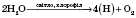
световая фаза
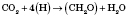
темновая фаза
В световой фазе фотосинтеза происходит поглощение света молекулами хлорофилла и трансформация энергии света в химическую энергию АТФ и восстановленного НАДФН (нікотинамідаденіндінуклеотидфосфат восстановлен). Эти процессы осуществляются белковыми комплексами, входящими в состав тилакоїдів хлоропластов.
Образованные в результате фотохимических реакций АТФ и НАДФН используются для осуществления реакций темновой фазы, в которой происходит восстановление молекул СО2 до молекулы углеводов (глюкозы). Существуют различные способы восстановления СО2, самый распространенный из них - цикл Кальвина.
Глюкоза, образовавшаяся во время цикла Кальвина, может потом расщепляться до пірувата, поступать в цикл Кребса. Растения используют глюкозу как источник энергии в ночное время и для других процессов, в которых необходимо быстрое ее получение (движения листьев у бальзамина, росянки). При этом кислород не выделяется, а поглощается, а углекислый газ, образовавшийся выделяется в окружающую среду.
Биосинтез белков
В клеточном ядре происходит транскрипция - синтез молекулы іРНК на матрице ДНК по принципу комплементарности. Поэтому молекула (первичный транскрипт), которая образуется при этом, содержит не только кодуючі (екзони), но и некодуючі (интроне) последовательности. Особен-ные ферменты ядра способны познавать интроне и вырезать их. Кроме вырезания интронов, іРНК подвергается в ядре и другим модификациям: к ее концам прикрепляются сигнальные последовательности нуклеотидов, которые отвечают за дальнейшее соединение іРНК с рибосомою и за ее транспорт из ядра. Зрелые молекулы іРНК распознаются особыми белками ядерных пор, которые способствуют их продвижению в цитоплазму с помощью активного транспорта. Попав в цитоплазму, іРНК связывается с рибосомальними субъединицами, будучи сигналом для их сбора в функционально активную рибосому.
Трансляция. В процессе трансляции нуклеотидная последовательность іРНК считывается группами по три нуклеотиды (такие триплеты называют кодонами), по мере того как рибосома перемещается вдоль молекулы іРНК. Каждая аминокислота соответствует определенному кодону.
Транспорт аминокислот к рибосомам обеспечивают тРНК. Для каждой аминокислоты есть специфическая тРНК.
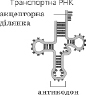
Транспортные РНК выполняют роль звеньев, которые связывают триплетний код, содержащийся в іРНК, и амінокислотну последовательность полипептидной цепи. Сравнительно небольшие молекулы тРНК содержат около 80 нуклеотидов. Все молекулы имеют схожую структуру: в каждой есть акцепторна участок, к которому присоединяется соответствующая аминокислота, участок, которая содержит антикодон, - последовательность из трех нуклеотидов, комплементарную кодону іРНК, который соответствует определенной амінокислоті.
Транспортная РНК с присоединенной к ней аминокислотой подходит к рибосомы и связывается с комплементарным антикодоном триплетом (кордоном) молекулы іРНК. Связывание происходит в строго определенном месте - на так называемой А-участке рибосомы. В этот момент на Р-участке (она находится рядом с А-участком) уже есть тРНК, которая удерживает конец растущей полипептидной цепи. Аминокислота, закрепленная на новоприбулій тРНК, образует пептидную связь с СООН-концевой аминокислотой полипептидной цепи, и тРНК, которая находилась до этого на Р-участке, отделяется от рибосомы и способна транспортировать другую такую же аминокислоту. Это приводит к перемещению тРНК (с которым теперь связаны аминокислоты белка, который синтезируется), которая осталась, на освобожденную Р-участок. Теперь А-участок доступна для прикрепления следующей молекулы тРНК, антикодон которой комплементарный кодону іРНК (последняя также переместилась в отношении А-участка на один триплет). Так продолжается до тех пор, пока в А-участке рибосомы не окажется кодон іРНК, который не кодирует никакой аминокислоты - стоп-кодон. В эукариот стоп-кодонами являются триплеты УАА, УАГ и УГА. В них нет комплементарного антикодону тРНК, отсутствие тРНК в А-участке вызывает отщепление полипептидной цепи от тРНК, находящейся в Р-участке. Трансляция прекращается.
Скорость сборки белка может быть увеличена, если синтез полипептидной цепи происходит на полірибосомальному комплексе (полісомі).