Раздел 1
БИОЛОГИЧЕСКИЕ ОСНОВЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
1.2. Молекулярно-генетический и
клеточный уровни организации жизни
1.2.2. Структурно-химическая и
функциональная организация клеток еукаріотичних
1.2.2.6. Органеллы цитоплазмы
Клеточные органеллы - дифференцированные
участка цитоплазмы, имеющие специфический молекулярный состав. Это сложные,
високовпорядковані биологические системы макромолекул, образующих определенную
пространственную структуру, способны к выполнению специальных клеточных функций.
Клетки животных содержат много внутриклеточных мембран. Поэтому почти половина
всего объема клеток заключена в отдельные внутриклеточные отсеки (компартменти),
что называются "органелами" (рис. 1.29). Другой внутриклеточный
пространство занят цитозолем.
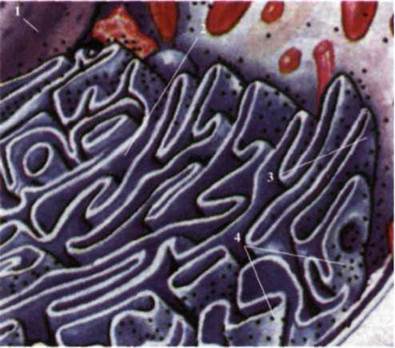
Рис. 1.29. Клетка эукариот:
1 - ядро; 2 - гладкая
эндоплазматическая сеть; С - зернистая эндоплазматическая сеть; 4 - рибосомы.
Классификация органелл. Клеточные
органеллы условно подразделяют на мембранные, которые окружены типичной біомембраною, и немембранні, что не
имеют такой оболочки.
Мембранные:
1) эндоплазматическая сеть: а) зернистая; б) гладкая; 2) комплекс Голги; 3)
лизосомы; 4) пероксисоми; 5) вакуоли; 6) митохондрии; 7) пластиди (только в
растительных клетках).
Немембранні: 1) рибосомы; 2) центріолі;
3) микротрубочки; 4) мікрофіламенти.
Согласно выполняемых функций
различают органеллы общего и специального назначения.
Органеллы общего назначения
встречаются во всех еукаріотичних клетках и относятся к общим структур.
Специальные органеллы характерны
только для определенного вида клеток, выполняющих специфическую функцию. Например, в
некоторых простейших - это жгутики, сократительная вакуоля, ундулююча мембрана. В
мышечных клетках - скоротливе волокно; нейроны имеют длинные отростки,
сперматозоид - акросому и т.д.
Эндоплазматическая сеть (ЭПС). ЭПС
обнаружена во всех еукаріотичних клетках, отсутствует только у прокариот, в
сперматозоидах и зрелых эритроцитах. ЭПС образована сеткой мембранных трубочек, цистерн и овальных везикул. ЭПС структурно
связана с оболочкой ядра (рис. 1.29). Различают два типа ЭПС: гладкую и
зернистую, хотя они структурно связаны между собой. Зернистая ЭПС на своей
поверхности содержит рибосомы, которых нет на поверхности гладкой ЭПС. ЭПС образует
сетку мембранных каналов, пронизывающих цитоплазму.
Эндоплазматическая сетка имеет значение в
процессах внутриклеточного обмена, поскольку увеличивает площадь внутренних
поверхностей клетки, разделяет ее на отсеки, отличающихся по физическим состоянием и химическим составом, обеспечивает
изоляцию ферментных систем, что, в свою очередь, необходимое для последовательного вступления
в согласованные реакции. Непосредственным продолжением эндоплазматической сетки является ядерная
мембрана, которая отделяет ядро от цитоплазмы, так и внешняя мембрана
(плазмолема), расположенная на периферии клетки. Мембранные системы очень лабильные
и могут меняться в зависимости от физиологического состояния клетки, характера
обмена, при росте и дифференцировании.
Цитоплазма клеток еукаріотичних
содержит мембранные слои, пузырьки, трубочки, отделяющие в совокупности
значительный внутриклеточное пространство. Мембраны ЭПС образуют непрерывные
структуры с наружной ядерной мембраной, они специализируются на синтезе и
транспорте липидов и мембранных белков. Зернистая ЭПС выглядит как система
плоских цистерн, внешняя сторона которых покрыта рибосомами, которые синтезируют белки.
Гладкая ЭПС, трубчатой строения, не имеет рибосом.
Гладкая ЭПС встречается в
клетках, которые выполняют секреторную функцию, мышечных и пигмент
них
клетках. Зернистая ЭПС хорошо развита в клетках печени, поджелудочной
железы, секреторных клетках, где образуется белковый секрет.
Общие функции ЭПС. Взаимосвязана система гладкой и зернистой ЭПС работает
согласовано и выполняет ряд общих интегральных функций: 1) мембраны ЭПС
отделяют свое специфическое содержание от цитозоля, образуют специальный
компартмент; 2) в матриксе ЭПС происходит накопление, сохранение и
модификация синтезированных веществ; 3) ЭПС является важной составляющей системы
внутриклеточных мембран, обеспечивает транспорт синтезированных веществ во внутренних
полостях или с помощью везикул в разные участки клеток;
4)
структура ЭПС образует мембранную поверхность внутри клетки, что важно
для многих метаболических реакций; 5) мембранная система пронизывает всю клетку и
выступает в качестве "внутреннего скелета".
Комплекс Голги. Комплекс Голги
(КГ), образованный комплексом из десятков уплощенных дискообразных мембранных
цистерн, мешочков, трубочек и везикул, в значительном количестве встречается в
секреторных клетках. Внутренний міжмембранний пространство заполнено матриксом,
содержит специальные ферменты.
Электронно-микроскопические исследования
позволили убедиться, что КГ построен из мембран и напоминает столбик из пустых
дисков, наложенных друг на друга (рис. 1.30). В его состав входит система
трубочек с пузырьками на концах. Комплекс Голги имеет две зоны: зону
формирование, куда поступает синтезированный материал из ЭПС с помощью
транспортных везикул, и зону созревания, где формируется секрет и зрелые
секреторные мешочки.
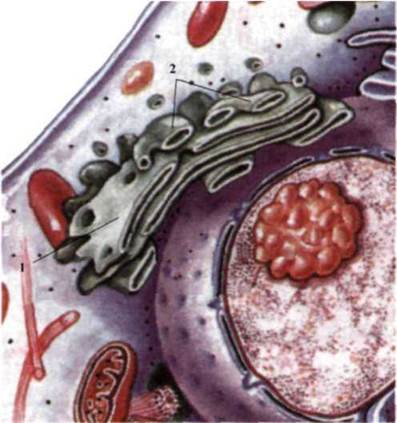
Рис. 1.30. Комплекс Голги:
1 - зона формирования; 2 - зона
созревания.
До зоны формирования поступают
синтезированные в ЭПС вещества, находящиеся в мембранных везикулах. Они
сливаются с мембраной КГ, и содержимое везикулы поступает внутрь комплекса.
Вещества обрабатываются ферментами, после этого снова упаковываются в везикулы и
переносятся в зону созревания.
В зоне созревания накапливается
"созревший секрет", что отделяется в виде секреторных пузырьков.
В этом компартменті образуются также лизосомы и пероксисоми.
Функции
комплекса Голги: 1) накопление и модификация синтезированных макромолекул; 2)
образование сложных секретов и секреторных везикул; 3) синтез и модификация
углеводов, образование гликопротеидов; 4) КГ играет важную роль в восстановлении
цитоплазматической мембраны путем образования мембранных везикул и следующего
слияние с клеточной оболочкой; 5) образование лизосом; 6) образование пероксисом.
Специальные функции комплекса
Голги: 1) формирование акросомы сперматозоида во время сперматогенеза; 2) вітелогенез - процесс синтеза и
формирование желтка в яйцеклетке.
Таким образом, является главным КГ
регулятором движения макромолекул в клетке, он собирает синтезированные белки, жиры,
углеводы, формирует транспортные везикулы и распределяет по клетке и за ее пределы.
Лизосомы - это небольшие (0,2-0,8
мкм), покрытые мембраной, круглые тельца (рис. 1.31). Встречаются они во всех
клетках растений и животных, могут локализоваться в любом месте клетки.
Содержимое лизосом составляют различные классы гидролитических ферментов, например,
протеазы, нуклеазы, липазы, фосфолипазы и др. Всего насчитывается до 40
различных ферментов.
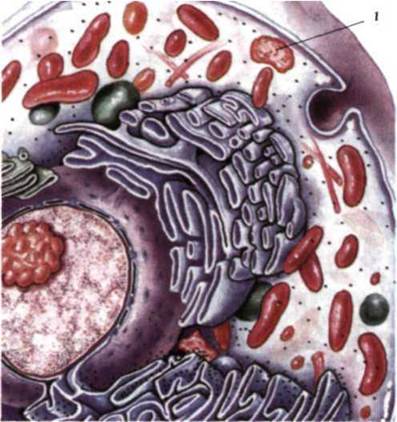
Рис. 1.31. Клетка эукариот:
1 - лізосома.
Эти ферменты разрушают крупные молекулы
сложных органических соединений, поступающих в клетки (белки, нуклеиновые кислоты,
полисахариды). В лизосомах подвергаются разрушению микроорганизмы и вирусы. Ферменты
лизосом переваривают разрушенные структуры или целые клетки. Эти процессы
называются аутофагією (от греч. αυτός- самый, φάγος- пожирание).
Лизосомы играют также существенную
роль в индивидуальном развитии организмов. Они разрушают временные органы
эмбрионов и личинок, например, жабры и хвост у головастиков лягушки, перепонки между
пальцами у эмбриона человека и др.
Каждая лізосома покрыта плотной
мембраной, изолирующей ферменты от цитоплазмы. Повреждение мембран лизосом и выход из них в цитоплазму
ферментов вызывает быстрое растворение (лизис) клетки.
Потеря лізосомами любой
ферментативной системы приводит к тяжелым патологических состояний целого
организма, наследственных болезней. Они получили название болезней накопления,
поскольку связанные с накоплением в клетках "непереваренных"
веществ, что мешает нормальному функционированию клетки. Эти болезни могут
проявляться развитием скелета, отдельных внутренних органов,
центральной нервной системы и др. С дефицитом лизосомных ферментов связывают
развитие атеросклероза, ожирения и других нарушений. С другой стороны, патологическая
активность лизосом может привести к разрушению жизненно важных структур.
Лизосомы разнообразные по своей
природой и могут образовываться различными путями.
В каждом случае формируются
морфологически разнообразные лизосомы, расщепляющих материал из разных источников. В
центре этих путей находится "промежуточный компартмент" - ендолізосома.
Процесс переваривания лизосомными
ферментами объектов, поступающих в клетку путем фагоцитоза, происходит в вакуолях, которые
называются фагосомами. Продукты переваривания попадают в цитоплазму, а
непереваренный материал остается в фагосомах и уменьшается в размерах. Такие
структуры называются остаточными тельцами. Они могут быть различной плотности
и размера.
Ендосоми лизосом могут сливаться с
внутренними структурами и разрушать их. В клетке при этом образуются большие
мешочки, покрытые общей мембраной, различной формы и плотности. Такие тельца
называются аутофагосомами.
Функции лизосом: 1) переваривание
веществ, поступающих в клетки из окружающего пространства (фагоцитоз), в частности,
таким способом организм борется с
микробами и вирусами; 2) переваривания внутриклеточных макромолекул,
выполнили свою функцию, и органелл (аутофагоцитоз); 3) переваривание погибших
клеток, или тех, что выполнили свою функцию; 4) рециклизация органических молекул -
расщепление использованных белков, а также углеводов, нуклеиновых кислот к
мономеров (аминокислот, моносахаридов, нуклеотидов) и повторное их использование
клеткой для синтеза новых молекул. Этим достигается экономичность
(многократность) использование внутренних молекул.
Пероксисоми. Пероксисоми - маленькие
сферические тельца, покрытые мембраной (рис. 1.32). Обнаруживаются почти у всех
клетках еукаріотів. их диаметр составляет 0,3-1,0 мкм, образуются в комплексе
Голги. Пероксисоми содержат в основном ферменты для разрушения пероксида водорода. Пероксид водорода, образующийся в результате
окисления некоторых органических веществ, является токсичным для клетки и поэтому немедленно
разрушается каталазой пероксисоми:

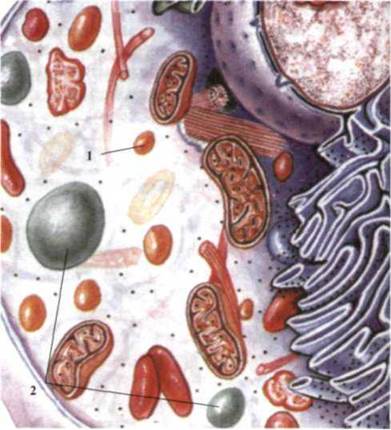
Рис. 1.32. Клетка эукариот:
1 - пероксисомы, 2 - вакуоли.
Пероксисоми участвуют в процессе
Р-окисления жирных кислот. До 50 % жирных кислот разрушаются в пероксисомах.
Они содержат также и другие окислительные ферменты.
Вакуоли. Вакуоли - это полости в
цитоплазме, окруженные мембраной и заполненные жидкостью (рис. 1.32). В еукаріотичних
клетках есть разные типы вакуолей. Вакуоли могут возникать из пузырьков, которые
отделяются от эндоплазматической сети, или комплекса Голги. Они
заполненные водным раствором органических и неорганических соединений, среди них -
продуктов обмена или пигментов. Функции вакуолей разнообразны: они поддерживают
тургорного давления, сохраняют питательные вещества и накапливают продукты обмена.
Сократительные вакуоли одноклеточных животных регулируют осмотическое давление в клетке,
участвуют в выведении продуктов обмена, а также способствуют поступлению в
клетку воды.
Митохондрии. Митохондрии (от греч. μίτος- нить, χονδράς - зернышко) - это органеллы, в которых
энергия химических связей органических веществ превращается в энергию
фосфатных связей АТФ. Митохондрии - довольно крупные овальные органеллы (0,2-2,0
мкм), покрыты двумя мембранами. Они встречаются почти во всех еукаріотичних
клетках, за исключением анаэробных простейших и эритроцитов. Митохондрии
хаотично распределены по цитоплазме, хотя чаще оказываются возле ядра или в
местах с высокими потребностями в энергии. В мышечных клетках они расположены между
міофібрилами. Органеллы могут изменять свою структуру и форму, способные
перемещаться внутри клетки. Количество митохондрий может меняться
в зависимости от активности клетки от нескольких десятков до нескольких тысяч.
Органела содержит внешнюю и
внутреннюю мембраны с узким пространством міжмембранним
(рис. 1.33). Внутренняя мембрана образует многочисленные выросты - кристи, окруженные матриксом, в котором находится много
ферменты, рибосомы, одна молекула ДНК. Митохондрии - органеллы размером с бактерию,
что используют энергию окисления для образования АТФ.
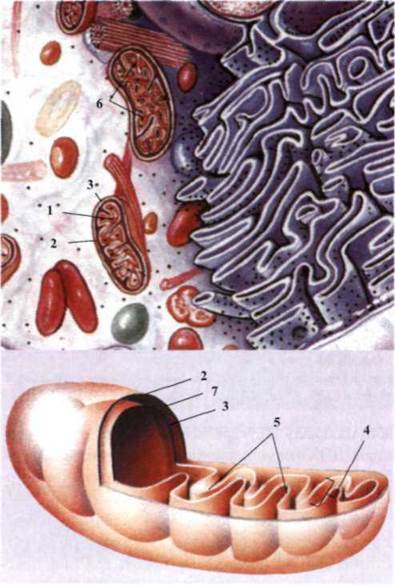
Рис. 1.33. Клетка эукариот:
1 - митохондрия; 2 - внешняя
мембрана; 3 - внутренняя мембрана; 4 - криста;
5 - матрикс; 6 - рибосома; 7 - міжмембранний
пространство.
Внешняя мембрана легко проницаема для
многих небольших молекул. Содержит ферменты, преобразующие вещества на
реакционноспособные субстраты, участвует в образовании міжмембранного пространства.
Внутренняя мембрана плохо проницаемая
для большинства веществ. Она образует выросты - кристи внутри матрикса. Эта
мембрана содержит ферменты, которые принимают участие в следующих важных процессах:
а) ферменты, катализирующие
окислительно-восстановительные реакции дыхательной цепи и транспорта электронов. В
результате образуется избыток Н+ в міжмембранному пространстве;
б) специфические транспортные белки
участвуют в образовании градиента Н+;
в) ферментный комплекс
АТФ-синтетазы, что синтезирует АТФ.
Міжмембранний пространство
используется для градиента ионов Н+ на внутренней мембране, что является
необходимым условием синтеза АТФ.
Матрикс - это пространство митохондрии,
ограниченное внутренней мембраной. Он образованный сотнями различных ферментов,
принимают участие в разрушении органических веществ до СО2 и Н2О.
При этом высвобождается энергия химических связей между атомами молекул
органических веществ и превращается в макроэргические связи АТФ. В матриксе
находятся рибосомы и молекула митохондриальной ДНК. Рибосомы митохондрий и
ДНК обеспечивают синтез необходимых органелі белков. Основная интегральная функция
митохондрий - преобразования энергии и образования АТФ, что содержит макроэргические
связи.
К митохондрии поступают разные
органические вещества, которые в матриксе окисляющиеся до простейших соединений (СО2
и Н2О). Внутренние мембраны имеют необходимый набор ферментов для
преобразование энергии химических связей, высвобождающуюся в энергию АТФ.
Пируват и жирные кислоты поступают в митохондрии и метаболизируются в цикле
лимонной кислоты, в котором образуется НАДН. Затем в процессе окислительного
фосфорилирования богатые на энергию электронов НАДН передаются на кислород по
помощью дыхательной цепи, что находится на внутренней мембране. При этом
в міжмембранному пространстве возникает высокая концентрация Н+. Это создает
электрохимический потенциал на внутренней мембране. Благодаря такому
хеміосмотичному механизма образуется АТФ. АТФ-синтетаза, что использует
энергию градиента Н+ из АДФ и неорганического фосфата, синтезирует АТФ. АДФ + Фн + энергия
градиента № → АТФ. Энергия АТФ используется
клеткой на движение, сокращение, синтез, транспорт и т.д.
Митохондрии размножаются путем
разделения. При делении клетки они более-менее равномерно распределяются между
дочерними клетками. Таким образом между митохондриями последовательных генераций клеток
осуществляется преемственность.
Особенности митохондрий, что указывают
на их сходство с прокаріотами, рассматривают как доказательство симбиотического
происхождение этой органеллы. Согласно такой гипотезе, некоторые аэробные прокариоты
проникли в большую анаэробную клетку. Возможно, сначала они вели паразитический
способ жизни. В дальнейшем партнеры этого сожительства в процессе эволюции приспособились
друг к другу и бывший "паразит" превратился в органелу,
необходимую для существования клетки. Но как органеллы предки митохондрии потеряли
часть своего генетического материала. В еукаріотичних клетках митохондриальная
ДНК кодирует лишь часть митохондриальных белков, большее же их количество
синтезируется вне митохондриями и связана с ядерной ДНК.
Пластиди. Пластиди - двомембранні
органеллы клеток растений и некоторых животных (жгутиковых). В клетках высших растений
различают три типа пластид: хлоропласты, хромопласти и лейкопласты.
Хлоропласты - окрашенные в зеленый
цвет благодаря пигмента хлорофилла. Между наружной и внутренней мембранами
хлоропластов является міжмембранний пространство шириной около 20-30 нм. Внутренняя
мембрана образует вгины - ламели и тилакоїди. Ламели имеют вид плоских
удлиненных складок, а тилакоїди - уплощенных вакуолей или мешочков. Ламели образуют сетку разветвленных
канальцев. Между ламелами размещены тилакоїди, собранные в виде стопки монет (граны). В тилакоїдах
находятся фотосинтетичні пигменты - хлорофилл, каротиноиды и ферменты, которые
нужны для осуществления различных биохимических процессов (рис. 1.34). В
матриксе пластид есть также собственный білоксинтезувальний аппарат (молекулы ДНК и
рибосомы). Основная функция хлоропластов - фотосинтез.
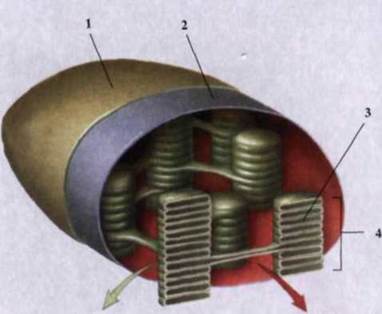
Рис. 1.34. Строение хлоропласта:
1 - наружная мембрана; 2 - внутренняя
мембрана; С - тилакоїд; 4 - грана.
Лейкопласты - бесцветные пластиди,
которые отличаются от хлоропластов отсутствием развитой ламелярной системы.
Они обеспечивают синтез и гидролиз крахмала и белков.
Хромопласти - пластиди, которые предоставляют
окраска (желтого, красного и др.) лепесткам, плодам, листьям. Окраска
хромопластів обусловливают пигменты - каротиноиды. Внутренняя мембранная система в
хромопластів отсутствует или образована единичными тилакоїдами.
Рибосомы. Рибосомы (от лат. ribes -
поток, струя и греч. σώμα - тело) - небольшие гранулоподібні
сферические тельца, размером от 15 до 35 нм. Рибосомы состоят из двух
субъединиц, расположенные в цитоплазматичному матриксе или связаны с мембранами
эндоплазматической сети (рис. 1.35).
Рис. 1.35. Клетка эукариот: 1 - рибосомы.
Субъединицы рибосом образуются в
ядерці, а потом через ядерные поры отдельно друг от друга поступают в
цитоплазми. их количество в цитоплазме зависит от синтетической активности
клетки и может составлять от сотни до нескольких тысяч на одну клетку, их функцией
является синтез белков. Наибольшее количество рибосом обнаружена в клетках, интенсивно
синтезируют белки. Эти органеллы встречаются также в матриксе мітохондріальному
и хлоропластах.
Рибосомы любых организмов - от
бактерий до млекопитающих - характеризуются сходством структуры и состава. Каждая
субъединица состоит из нескольких разновидностей молекул рРНК и десятков разновидностей
белков, примерно в одинаковой пропорции. Маленькая и большая субъединицы находятся
в цитоплазме отдельно друг от друга, пока не участвуют в белковом синтезе.
Они объединяются друг с другом и с молекулой ІРНК по необходимости синтеза и
опять разъединяются с прекращением процесса.
Если с одной молекулой ІРНК
соединяются несколько рибосом, то образуются полісоми, содержащих от 5 до 70
рибосом.
Клеточный центр (центросома) -
органела, состоящий из двух мелких образований: центріоль и лучевой сферы
вокруг них (рис. 1.36). С помощью электронного микроскопа установлено, что
каждая центріоля - это цилиндрическое тельце длиной 0,3-0,5 мкм и диаметром
около 0,15 мкм. Стенки цилиндра состоят из 9 пар параллельно расположенных
микротрубочек, которые образованы белками. Цент - ріолі размещаются перпендикулярно
одна к одной.
Рис. 1.36. Клетка эукариот: 1 -
центроєома.
Клеточный центр иногда занимает
геометрический центр клетки (отсюда название органеллы). Чаще же он отжатый
ядром или включениями к периферии, но обязательно располагается вблизи ядра
на одной оси с центром ядра и клетки. Активная роль клеточного центра
оказывается при делении клетки. Центріолі удваиваются и расходясь в
противоположные стороны, формируют полюса клетки, что делится. Указанные структуры
образуют веретено деления.
Центріолі участвуют в образовании
микротрубочек цитоскелета. Они также формируют базальне тело, что лежит в
основе жгутиков.
Микротрубочки и мікрофіламенти.
Микротрубочки и мікрофіламенти - это немембранні органеллы, которые построены из
сократительных белков (тубулина, актина, миозина и др). Микротрубочки (рис. 1.37)
цилиндрической формы, полые, диаметром 10-25 нм. Они участвуют в
формировании веретена деления, во внутриклеточном транспорте веществ, входящих
в состав ресничек, жгутиков, центріоль.
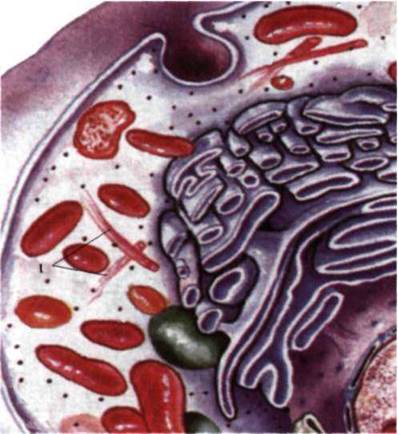
Рис. 1.37. Клетка эукариот: 1 -
микротрубочки.
Мікрофіламенти причастны к
формирование цитоскелета клетки. Расположены эти органеллы под плазматичною
мембраной. Пучки мікрофіламентів одним концом прикрепляются к мембране, а
другим - в разных органелл, молекул биополимеров. Они участвуют в изменении формы
клетки, например, во время ее движения. В мышечных клетках пучки
мікрофіламентів размещены вдоль их оси (волокна актина
и миозина).