Раздел 1
БИОЛОГИЧЕСКИЕ ОСНОВЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
1.3. Онтогенетический уровень
организации жизни
1.3.3. Биология индивидуального
развития
1.3.3.3. Проблема детерминации и
взаимодействия бластомеров
Основоположником экспериментальной
эмбриологии стал в конце XIX века немецкий зоолог В. Ру. Он попытался
выяснить, на какой стадии развития зародыша определяется судьба его частей. Или
может, например, каждый из двух первых бластомеров развиться в целый организм?
С этой целью В. Ру один из двух бластомеров лягушки разрушал раскаленной иглой.
С незруйнованого (целого)
бластомера развился неполноценный зародыш. Это дало повод В. Ру дойти
неверному выводу о том, что уже на ранних стадиях эмбрионального развития
судьба отдельных бластомеров полностью определена. Здесь нужно отметить, что в ходе
эксперимента ученый оставлял поврежден бластомер наряду с целым (невредимым).
Позже другие ученые-экспериментаторы провели эксперимент иначе. Они
забирали поврежден бластомер от целого и тогда наблюдали, как
из последнего развивался полноценный зародыш. Ученик В. Ру Г. Дріш показал: если яйца морского ежа поместить в
морскую воду, лишенную кальция, то в зиготі, что начала делиться, бластомери
распадаются и затем каждый из них при благоприятных условиях способен сформировать
полноценный зародыш. Такое явление, когда изолированный бластомер может развиться в
полноценный организм, получило название тотіпотентність (рівноспадковість), а такие
бластомери назвали тотіпотентними (рис. 1.160). В дальнейшем, опять же
экспериментальным путем было установлено, что у тритона сохраняется такая
тотіпотентність до стадии 16 бластомеров, у кролика - к стадии 4 бластомеров.
О существовании подобного явления в бластомеров человека указывают случаи рождения
двух, трех, четырех и даже семи монозиготних близнецов.

Рис. 1.160. Бластомери тотіпотентні.
После образования бластули зародышевые
клетки теряют тотіпотентність. Начинается дифференциация, то есть формирование
разнообразных структур и частей тела (а впоследствии - тканей и органов) с относительно
однородного материала зародыша. Но выяснилось, что, несмотря на потерю
тотипотентности и дифференцировки, клетки полностью сохраняют генетическую
информацию. Это показали в своих экспериментах американский ботаник Стюард на
моркови и английский зоолог Д. Гердон (1964-1966), который пересаживал ядра с
клеток эпителия кожи и кишок головастиков в яйцеклетки, из которых их собственные ядра
удалялись. Многие из таких клеток развились, образовав головастиков и даже
половозрелых лягушек (рис. 1.161).
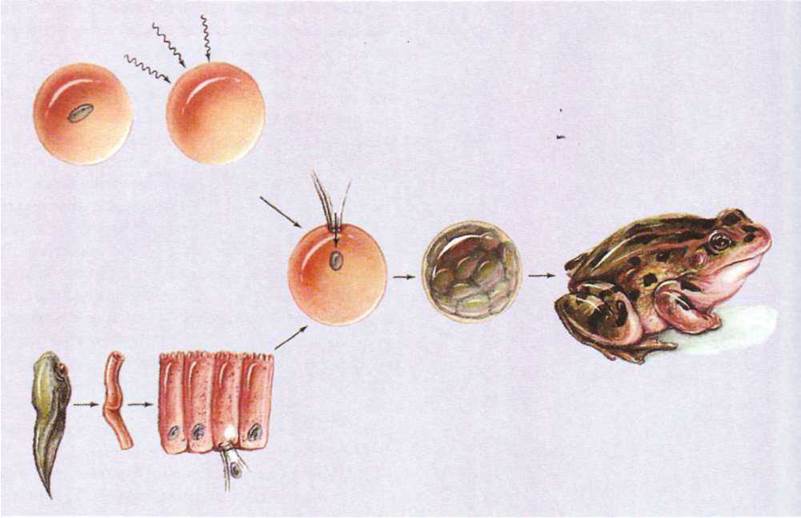
Рис. 1.161. Ядерная трансплантация
(опыт Д. Гердона).
Таким образом, можно прийти к выводу,
что любая соматическая клетка, с одной стороны, представляет собой дифференцированную
часть организма и выполняет узко специализированные функции, а с другой стороны,
удерживает в себе генотип всего организма. Тогда следует вопрос : если генотип в
всех клеток одинаков, а разные клетки имеют неодинаковую структуру и входят в
состав органов, отличающихся друг от друга по морфологии и по функциям,
то чем обусловлено дифференцировки во время развития?
Чтобы разобраться в этом вопросе,
вернемся снова к эксперименту В. Ру с двумя бластомерами зародыша лягушки. Ру убивал один из
бластомеров, но оставлял его рядом с целым, невредимым, и наблюдал
неполноценный его развитие. То есть тотіпотентність не проявлялась. Почему? А
потому, что поврежденный бластомер влиял и препятствовал нормальному развитию второго целого бластомера.
Так почти 100 лет назад была обнаружена и доказана взаимодействие частей организма в
процессе развития.
Позже это явление взаимного влияния
одних частей развивающегося организма на другие получило название эмбриональной
индукции.
Первым, кто убедительно доказал
взаимодействие частей зародыша на примере формирования глаза, был в начале XX века
немецкий эмбриолог Г. Шпеман (1864-1941). Он автор оригинальной теории
индивидуального развития - теории организационных центров.
Ранняя закладка глаза представляет собой
выпячивание ткани промежуточного мозга в так называемый глазной пузырь. Он растет в
направлению к кожной эктодермы, из которой на месте их сближения образуется
хрусталик. В эксперименте было показано, что если на одной стороне зародыша
удалить закладку глазного пузыря, то на этой стороне хрусталик не формируется. И,
наоборот, если закладка глазного пузыря пересадить под кожу в другом месте головы
или туловища, то здесь на границе эктодермы возникает хрусталик. Он в свою очередь,
повлияет на формирование глазной впадины. Таким образом, развитие одного органа обуславливает формирование следующих.
Дальнейшими исследованиями было
раскрыты новые звенья воздействия между частями зародыша. Г. Шпеманом было доказано, что
при нормальном развитии спинная ектодерма гаструли превращается в зачаток нервной трубки, но если ее
пересадить, например, на брюшную часть зародыша, то из нее образуется кожа
живота. И, наоборот, если кожную ектодерму живота пересадить на спинную часть
зародыша, то из нее образуется нервная трубка (рис. 1.162).
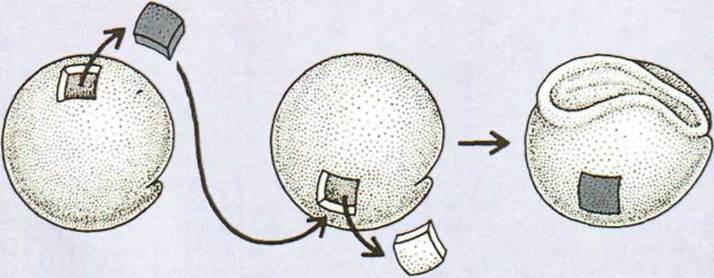
Рис. 1.162. Влияние между частями
зародыша (опыт Г. Шпемана).
Это свидетельствует о том, что на стадии
ранней гаструли судьба зачатков еще не полностью определена и возможно
переопределение дальнейшего пути развития. Эта стадия эмбрионального развития
получила название стадии лабильного дифференцировки.
Почему же ектодерма спинной части
зародыша дает начало нервной трубке со всеми ее производными? Экспериментально
было установлено, что если на ранней стадии удалить участок верхней
"губы бластопора, которая и прилегает к эктодермы на спинной стороне зародыша,
то нервная трубка совсем не образуется. И, наоборот, если верхнюю
"губу" бластопора пересадить под кожу в любом участке гаструли,
то из окружающих клеток формируется нервная трубка. И даже если пересадить
дополнительную верхнюю "губу" бластопора, то образуется дополнительная нервная
трубка.
Так, Г. Шпеман пришел к выводу, что
верхняя "губа бластопора влияет на клетки эктодермы, направляя их
развитие на формирование нервной трубки. Впоследствии были обнаружены другие участки тела
зародыша с аналогичным воздействием на его части. Эти участки были названы
организационными центрами, а их индуцированный влияние - организаторами (или
индукторами).
Реализация индукции возможна только
при условии, что клетки способны воспринимать воздействие, то есть являются компетентными. Компетенция
клеток может меняться во времени (возникать и исчезать). На более поздних стадиях
развития, когда дифференциация уже началась, пересадка этого участка в другую
часть зародыша не меняет его развитие, ибо он уже полностью определен. Это
явление независимой, стабильной дифференциации. В развитии органов наблюдается
переход от зависимого к независимому дифференциации.
Дальнейшие исследования проблемы
эмбриональной индукции шли в направлении определения природы индукторов.
Еще в 30-е годы XX в. американский
ученый Ч. Чайлд выдвинул предположение о градиент физиологической активности
организма зародыша. Было показано, что интенсивность окислительных и других
метаболических процессов постепенно тормозится в направлении от главного отдела к
хвостового. Согласно этим данным, развитие органов в той или иной части
зародыша определяется интенсивностью метаболизма. Данные согласуются с положениями
школы Шпемана: действительно, наиболее интенсивный обмен веществ в гаструли
происходит в участке верхней "губы бластопора, то есть в месте
образование нервной трубки.
X. Тидеман в 1970 году провел
исследования, которые пказали роль взаимоотношений органов зародыша на молекулярном
уровне. Он выделил из зародышей цыплят индукторы белковой природы. Один из них
индуцирует структуры нервной системы и органов чувств, другой - мышечную систему.
Ученый доказал, что белки-индукторы есть уже в незаплідненому яйце, но они
находятся в неактивном состоянии из-за присутствия ингибиторов, которые сдерживают их
действие. Во время эмбрионального развития изменяется ионный состав цитоплазмы и
внешней среды, которая окружает клетку. Это приводит к выходу ингибиторов
из клетки и этим инактивация из индукторов снимается. В результате этого
индукторы входят в ядро и активируют соответствующие гены, что, в свою очередь, способствует
дифференцировке клеток и формированию органов и систем.